|
|


|
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
简介(Brief Introduction) |
|
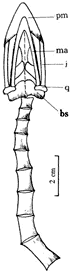
[A] 这一晚白垩世的鸟化石最突出的特征是有一长达305 mm、由20余枚尾椎骨组成的长尾. 尾椎骨保存19枚,前部椎体较短,具短的椎体横突,双凹型,但尾椎两侧未见羽毛印痕保存(图1,图2). 始祖鸟一百多年来一直吸引着世人的关注,尤其它那独特的长尾,是鸟类起源于爬行动物的显著性状之一[5,6]. 现在记述的雁荡鸟,是迄今为止除始祖鸟外,中生代所发现的完整鸟化石中唯一具有如此长尾的鸟化石. 我们同时也注意到在蒙古晚白垩世发现的Gobipteryx minuta和Mononykus olecranus两种鸟类化石[7,8]. 前者仅有部分头骨,从形态看,它与本文记述的雁荡鸟仅有两点相似:一是颌已无齿,二是头骨骨片较薄. Mononykus形态比较特殊,颌具带锯齿的锐利牙齿,前肢特化,很短,腰带完全与兽脚类恐龙一样,故至今极少有人承认它是鸟类,我们也认为它是一小型兽脚类恐龙,雁荡鸟很难与其对比. 雁荡鸟与始祖鸟及孔子鸟有着较多可相比之处[9,10],与中国其它中生代鸟类对比,不仅因它有一长尾,就是其它构造亦相差甚远.
图1 长尾雁荡鸟M1326骨骼图
图2 长尾雁荡鸟(新属、新种)Yandangornis logicaudus gen.et sp.nov.(M1326) 雁荡鸟头骨全长约50 mm,后部最宽约20 mm,头骨腹侧埋藏,故仅能观察到腹面的一些骨骼(图3). 吻较短,前颌骨发育,无齿而具角质喙与孔子鸟(Confuciusornis)是一致的[11]. 前颌骨较短和宽,无齿,眼前孔较大,颧弓也很扩展,下颌与上颌咬合,下颌窄薄,腹缘平,看不出有下颌孔. 方骨仅左侧者能看到,但保存不全,犁骨和颚骨观察不清,翼骨和基蝶骨亦仅保存部分骨片,枕孔的位置因挤压不能确定.
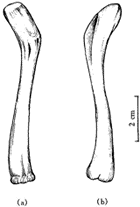
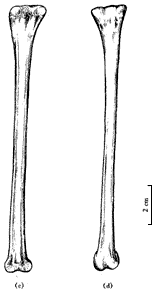
图3 长尾雁荡鸟头骨的腹视 颈长约80 mm,颈椎骨9枚,双凹型,前部椎体较短宽,后部椎体较长,颈肋已完全退化. 雁荡鸟与始祖鸟(Archaeopteryx)比较,其前肢构造很接近[12],特别是保存很好的一对肱骨(图4),近端构造简单,肱骨头小,但近端扩展部分比较长,没有气窝和顶沟等构造,已能看出骨体呈“S”形弯曲,肱骨远端内外关节髁已很分明,内髁大,已具有浅的鹰咀窝,较始祖鸟进步,与孔子鸟相似. 尺桡骨远端保存不全,桡骨直,较尺骨细,尺骨弯曲,近端已有短的鹰咀突和浅的杯状凹. 腕骨和掌骨缺失,至少有两个游离的指骨被保存下来,爪很小.
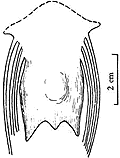
图4 长尾雁荡鸟左肱骨 胸骨长50 mm(图5),前部两侧有斜侧突,未见龙骨突,但胸骨中后部有明显的膨胀部,胸骨后部也有两个后侧突和较短的后突. 它与孔子鸟的胸骨相同之处是都为宽而大的板状,但孔子鸟的胸骨还没有前侧突. 另外,其腹侧还有腹肋残存,这又与始祖鸟、孔子鸟、中国鸟(Sinornis)[13]等相似.
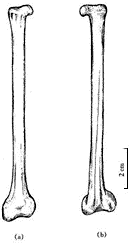
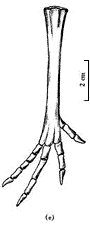
图5 长尾雁荡鸟胸骨腹视 雁荡鸟的肩带因挤压观察不清,在肱骨近端后缘仅能看到部分乌喙骨,它与胸骨接触的远端相当宽,推测不会太长,与一般中生代,尤其白垩纪鸟类乌喙骨的形态是一致的. 另外,保存了一部分叉骨,可能是左叉骨支的端部或是缝合部. 肩胛骨仅观察到右侧远端的一部分,它贴近胸骨的外边缘向后延伸,比较直和长. 腰带各骨仅能见到耻骨,它没有前突,其远端较细不扩展,很明显向后下方倾斜,坐骨和肠骨压在背侧看不清. 从耻骨的状况分析,三块骨骼不愈合. 肋骨细长,腹肋交叉排列很清楚. 后肢骨骼保存很好(图6),股骨较长而直,远端两关节髁很发育,向前突出,远远超出骨体之外,外髁较内髁大,髁间沟宽,外髁的外侧凹已很清楚,股骨全长约106 mm. 胫跗骨是长骨中最强壮的骨骼,近端与股骨关节的顶部扩展,已有明显的胫骨嵴,远端没有近端扩展,内、外关节髁较大,末端圆,胫跗骨全长132 mm. 腓骨仅能看到右侧者,比较细和短,其长不超过胫跗骨之半. 跗跖骨比较短,但粗壮,全长70 mm,仅为胫跗骨全长之半稍多一点,较股骨也短. 跗跖骨远端基本愈合,近端与胫跗骨关节的顶面扩展,已有两个浅杯状凹,跗跖骨远端第2,3,4趾骨滑车长短相差不大,第1趾骨(拇趾)与其它三趾相对,位置较高,有一小而不钩曲的爪,第2趾骨爪也比较弱,且不甚弯曲,较第1趾爪大,与中国早白垩世鸟类一般均具有大而弯曲的脚爪恰成鲜明对比,而和鱼鸟(Ichthyornis)很接近. 当然,雁荡鸟并不是水鸟.
图6 长尾雁荡鸟后肢 在研究过程中,我们注意到南美晚白垩世的几件原始鸟类化石,Patagopteryx, Neuquenornis, Aluarezsaurus[14],特别是最近发表的发现于马达加斯加岛晚白垩世的Rahona ostromi[15]. 前两者大小似鸡,前肢特别是肱骨缩短,而且Patagopteryx的颈椎已是明显的异凹型(heterocoelous),尾巴也明显缩短,与雁荡鸟不同;相似之处为眼孔大,腰带骨不愈合,后肢长短比例相似及趾爪不甚钩曲等. 有意义的是,雁荡鸟与Aluarezsaurus和Rahona有较多的相似性,如前肢不缩短,都具一较长的尾巴及后肢强壮,股骨与胫跗骨的比例相近等;但也有显著差异,如Aluarezsaurus和Rahona较雁荡鸟个体大,特别是雁荡鸟尺骨与股骨的比例与Rahona相反,股骨长于尺骨,Rahona的第二趾爪特别强大等. 尽管雁荡鸟与上述晚白垩世的鸟类有某些差异,但相似之处是明显的. 联系到我国南方这一时期的其它脊椎动物(如鱼、恐龙)与南美的相似性,雁荡鸟的发现似乎进一步提供了信息. 绝大多数早白垩世鸟类为适应飞行具有比较进步的前肢和比较后进的腰带及后肢,而本文所记述的晚白垩世特殊鸟类雁荡鸟却迥然不同. 这一不同寻常的鸟类给我们带来哪些关于鸟类及其演化的信息呢? 首先,本文讨论的雁荡鸟,国内外不少学者参观过,意见不一,有的认为可能是小型恐龙. 我们研究认为它是一原始的特殊鸟类,其依据是:(1) 头骨骨片薄,没有任何纹绉构造,下颌窄细;(2) 胸骨大,具前后侧突和后突;(3) 肱骨近端扩展部特别长,骨体具“S”形,远端内髁较外髁大;(4) 股骨第四转子退化,远端具较大的髁间凹;(5) 胫跗骨的胫嵴发达,腓骨细短;(6) 耻骨明显向后倾斜,而且耻骨没有前突;(7) 尾椎骨没有脉胍,仅前部几个尾椎骨有短的横突. 以上各点皆为鸟类特征,恐龙是不具备的. 雁荡鸟在鸟类系统分类中的位置,依据其长尾,前肢以及腹肋等特征,无疑应归入蜥鸟亚纲,我们也就是依此而将它放入该亚纲的. 但是,若从头骨和后肢以及胸骨的性状看,又与今鸟亚纲(Ornithurae)有比较密切的关系[16]. 已知中生代鸟类中,除始祖鸟、孔子鸟等晚侏罗世的原始鸟类还具有前肢的腕骨、掌骨和指骨等不愈合的特征外,其它白垩纪鸟类的前肢都较进步,各骨都趋于愈合,或者已基本愈合. 另外,尾椎亦绝大多数缩短,愈合为一原始的尾综骨,既使有极少数鸟尾椎不愈合也大大缩短了,如北美的黄昏鸟(Hesperornis)和蒙古的戈壁鸟等. 很显然,雁荡鸟是晚侏罗世蜥鸟亚纲成员的直接后裔. 其次,从适应功能的变化分析,雁荡鸟较侏罗纪原始鸟类出现晚几千万年,但前肢构造没有明显的进化,也没有突出的缩短,同时,其游离指爪很小,脚爪也不甚钩曲,显示出它已失去其祖先攀缘树木的能力,也就丧失了在树间滑翔的可能,再加上一条沉重的长尾,更无力飞离地面. 由于这种形态构造上的保守性,使之不可能向进步的飞翔方向发展,而只能在灌木丛中或河湖边缘的陆地上生活. 另外,从生态习性看,雁荡鸟的后肢强壮,各部构造相对进步,但它的趾骨较短,爪小,跗跖骨相对短而宽,胫跗骨长等,显示出它是适应地栖生活的原始鸟类. 因此,在生态习性上已与其发端于树栖生活的祖先有极大差异,这可能是由于它的遗传因子中,没有获得飞行能力的遗传密码. 从晚侏罗世到晚白垩世的六千余万年的地史时期(相当于整个新生代的历史)中,地壳经历过多次变化,特别是植被的演化,而雁荡鸟却摆脱不掉长尾巴的沉重负但,只能向着适应地栖生活的方向演化,它的前肢可能还会逐渐退化,它可能是后期陆生鸟类的最早代表. 既使是这样,雁荡鸟在与同时代的其它比较进步鸟类的生存竞争中,注定是弱者. 因为它不像鱼鸟那样既有飞翔能力,也适应地栖生活;更不具备黄昏鸟潜水的技能,以适于觅取水中食物. 这一晚白垩世早期原始鸟类的出现,使原本就复杂的有关鸟类的起源和进化更加复杂. 当然,这仍然显现出鸟类自爬行动物分化出不久即分为两大支系演化的事实,只是用传统进化理论不好解释它与比它早出现而又较它进步的鸟类之间的关系,因而又增加了鸟类进化过程的复杂性. 任何一类有生命的物种其演化过程绝不是线性的,而是曲折复杂的,人们尚需有更多的资料去认识,生物进化理论要不断地丰富和补充. 这里还有热河生物群义县组的两种著名脊椎动物化石要提一下,即中华鸟龙(Sinosouropteryx)[17]和原始祖鸟(Protarchaeopteryx)[18]. 这两种动物最初的鉴定者将它们都归属于最原始的鸟类,而前者一公布,几乎所有古生物学家都认为是恐龙,确实其形态与鸟类相距太远了,最近被陈丕基等正式修定归属于美颌龙科(Compsognathidae)[19]. 后者,最近又被鉴定者自己修定为恐龙了,而且又订了一新属种邹氏尾羽龙(Caudipteryx zoui)[20]. 它们确实与中华鸟龙不同,不但形态构造不同,而且其前肢和尾部都有羽毛状物,头高而短,颌具齿,前肢缩短,后肢强壮,但跖骨不但不愈合,而且彼此分离,趾骨很短,有向跖行性演化的趋势. 由于构造特殊,再加上特殊的皮肤覆盖物,其分类系统实难确定. 它们与雁荡鸟相似之处只是颈和尾都较长,其它构造绝然不同. 依据以上分析和讨论,雁荡鸟区别于已知的白垩纪其它鸟类,以此建立一个新目,雁荡鸟目(Yandangithformes ord.nov.),并归于蜥鸟亚纲. 参 考 文 献 [1] 侯连海. 中国中生代鸟类. 台北:台湾省立凤凰谷鸟园,1997. 201~207 |
|
化石写真(Fossil Pic.) |
|
|
|
复原图(Restitute Pic.) |
|
|
|
|
|
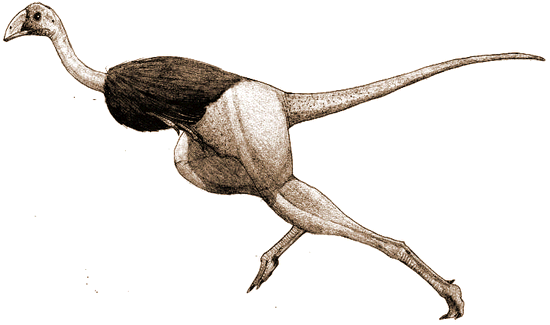
本站复原图(保留版权)(Copyrighted Restitute Pic.) |
|
|